

Photo by William Milliken, © RBG Kew (CC BY 3.0).

Photo by William Milliken, © RBG Kew (CC BY 3.0).
Tall: Eriophorum angustifolium. Tussock: Eriophorum vaginatum.
Tall: Least concern (IUCN; assessed March 4, 2013; stable population). Tussock: Least concern (IUCN; assessed January 10, 2015; unknown population stability).
Tall: 567 million base pairs. Tussock: 460 million base pairs.
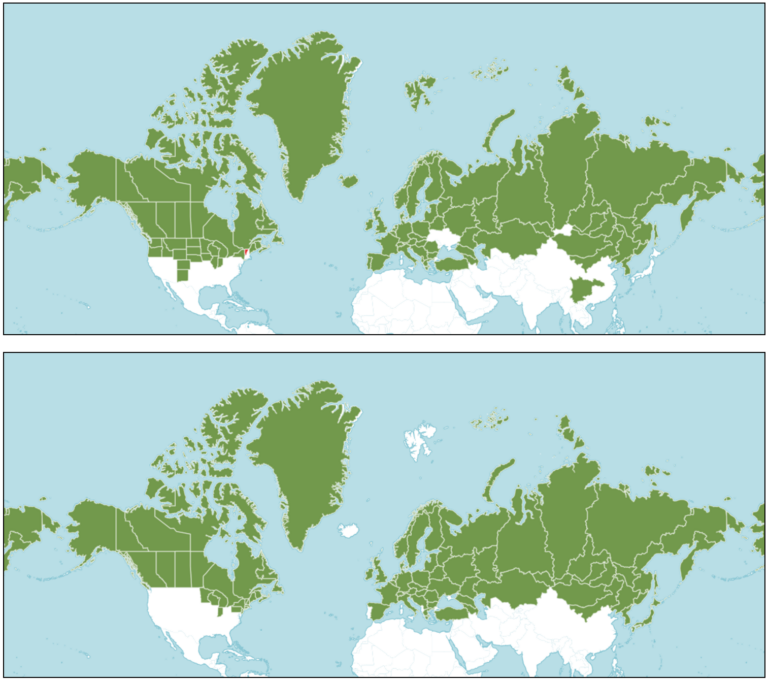
The sobriquet ‘cottongrass’ (vascular plants of the genus Eriophorum) describes the trait for which they are best known. The still-attached seeds of the plants give the appearance of tufts of cotton, reminiscent of the ‘true’ cotton plants in the genus Gossypium. With a secondary use as insulation for the plant, aiding survival in the frigid airs of northern latitudes, the primary function of the feathery tufts is to assist in seed dispersal, by catching the Fall wind and riding the flurries to parts unknown. When alighting on suitable substrate, the life history of cottongrasses begins anew, and germination begins in the following summer. For both of our cottongrass species, preferred substrate in the Arctic includes peaty and acidic soils in and around wetlands, amidst Carex species (another family of ‘sedges’) and other cottongrasses (such as E. scheuchzeri). However, the tussock cottongrass can also be found in small clumps, or hummocks (aka tussocks), and may be less tolerant of standing water than the tall cottongrass (and E. scheuchzeri). Concomitantly, the tall cottongrass fares better with where oxygen levels in soil are limiting, likely an adaptation to anoxic conditions of flooded peaty wetlands. In fact, in comparison to the tussock cottongrass, the tall cottongrass shows specific adaptations to existing in standing water, allowing both species to co-exist with reduced interspecific competition for resources. At the height of the Arctic summer with extended exposure to sunlight, cottongrasses may shut down photosynthesis, thus restricting growth, to limit the production of damaging ‘free radicals’ (highly reactive elements or compounds), as the tall cottongrass does in elevated Alpine habitat. Upon growth, Eriophorum species provide sustenance to a variety of animal life, including some terrestrial icons of the Arctic (caribou, muskoxen, and even grizzly bears), alongside lemmings, voles, geese, ptarmigans and perching birds (passerines), and various insect larvae. Their general high tolerance to seasonal extremes, and high levels of adaptability (via plasticity of developmental responses or heritable changes in genomes), have made cottongrasses, including the two highlighted here, very successful resident of peatlands, wetlands, and alpine regions throughout most of the northern hemisphere (see maps).
Qulliqs (or Kudliks) are traditional stone lamps used by Inuit which are dependent upon the use of earthbound (and seabound) biological resources for their use. The fuel for the lamp is usually seal blubber, but the wick with which to ignite and maintain a living light from the lamp is often the tufty feathers of cottongrasses. AINA staff are sometimes fortunate enough to see qulliqs in action during Arctic-held workshops, where they play a crucial part of blessing ceremonies conducted by community elders.
Cottongrass also has a history of use in traditional medicine. The feathery heads of cottongrasses have been used to heal navels of newborn infants, post parturition, by their being placed in the navel area sometimes with a little charcoal. Testimony by school children in Iqaluit also speak to their effectiveness as a remedy for sore throats. Raw stems have also been used a health restorative or as a medicinal drug for ailments.
Materially, all aspects of the plant have found utility in Inuit societies. The stems and roots of Eriophorum species provide a significant vegetative component to some Inuit diets. They can be eaten raw, boiled, cooked with seal blubber, or stored in seal blubber for later consumption. The plant fibres, derived from stems and/or leaves, have been used to add composition to mats, rugs, bedding and clothing, including using stems for weaving.
The IUCN has only assessed the conservation status of the tall cottongrass for its European distribution. Thus, from a global conservation standpoint, this species is data depauperate for its entire range, although there seems to be no real effort to delve further into what seems to be a common and widespread plant. The same status is afforded the tussock cottongrass. It seems that these widespread and foundational species are resilient enough to survive most threats. However, their usefulness in a conservation context might be more in how they respond to changes wrought by climate change in the Arctic, and how this may have broader and cascading impacts on the terrestrial Arctic ecosystem and its inhabitants in the future. The tussock cottongrass, in particular, being a foundational species of High Arctic tundra, has been used to experimentally test aspects of climatic upheaval in a warmer future. A warming of merely 1 degree centigrade can delay the timing of leaf ageing (senescence), extending the growing season. However, this may be offset by drying soil, which may reduce productivity (growth). However, not all ecotypes of the tussock cottongrass reacted equally, suggesting some inherent differences in populations to adapt to a warming climate. Experimental evidence suggests the more southerly ecotypes might be able to take advantage and subsequently move northwards at the expense of more northerly ecotypes so long as damp soils persist, as in one experiment the southerly populations were more prone to the negative impacts of dehydration of the soil than the northern ecotype. In European alpine regions, the tall cottongrass ‘rhizosphere’ (the soil root-fungal interface) has been associated with elevated levels of methanotrophic activity and thus reduced emissions of the greenhouse gas methane.
Finally, wildfires are strongly correlated with a warming and drying terrestrial landscape. Cottongrasses burn and are associated with fire risk. However, cottongrasses show some resistance to the effects of fire and post-fire recovery of tussock cottongrass may aid subsequent germination of seed sets of other plants. That said, if cottongrasses expand their range, especially southerly populations, then the risks of fire in the High Arctic may also increase in frequency, especially if the soil dries beyond a fire risk threshold.
Sedges are an incredibly successful group of vascular plants. Eriophorum sedges are no exception. Our subjects in this article, although not threatened with any existential crisis (barring any novel catastrophic agent appearing on the horizon such as disease), may provide invaluable insight on the adaptability and perseverance of what are called ecological generalists – those species that can survive, and even thrive, in different environments. In such an event, it’s a nice revelation that both the tall and the tussock cottongrass have chromosomal-level whole genome sequences declared recently, so that research into their respective biological differences (and similarities) can forge ahead. For plants, chromosomal-level WGS reveals the DNA sequence of each individual nuclear chromosome, but also the sequences of the organellar genomes – those of the mitochondria (‘powerhouses’ of the cell, occurring in animals, plants and fungi), and of the chloroplast (found only in plants, and which perform photosynthesis, the chemical reaction that manufactures chemical energy stores from sunlight, carbon dioxide and water; and upon which we all ultimately depend).
Both species have relatively small genomes (< 0.6 gigabases), if compared with a human being (3.42 gigabases), but the tall cottongrass (567 megabases) is near the average for vascular plants of 588 megabases. The real genomic leviathans of the plant kingdom are the gymnosperms (conifers, cycads, gynkos, etc) with an average of 15.68 gigabases! However, size isn’t everything and not a lot is known about why genomes vary so much in size even among species. Some species may even have whole genomes duplicated or reduced, or a combination thereof, differentiating distinct populations or subspecies, such as some Ambystoma mole salamanders. The duplication of whole genomes is relatively common in plant evolution and has occurred in the history of salmonid fishes and other cold-blooded vertebrates. Why some groups can undergo such whole genomic revolutions, but in others it is entirely absent or rare is (e.g., mammals) is a question for another day.
Some comparative genomics studies have started to investigate what parts of genomes are essential (‘conserved’), what parts are free to burgeon, diversify, or expand, according to the whims of particular evolutionary drivers; or if any of this genomic matter is, what used to be termed, ‘junk DNA’, with no palpable use beyond accretion during evolutionary descent from past generations with too high a cost of ditching it? This question has now begun to be talked about in Arctic flora. Initial work seems to point the finger at the presence (or absence) of repeated DNA sequence elements (called ‘motifs’) at the ends of chromosomes (called telomeres). These “DNA caps” seem to serve the function of a protective buffer to ward off continual chromosomal degradation (basically wear-and-tear) in plants that have extremely short growing seasons during which they rapidly make copies of genomes for new cell production and tissue growth. It is conceivable that differences in subtle growth requirements and tolerances between the tall and tussock cottongrass accounts for the near 100 million base pair differences (around 20 %) in size of their respective genomes, despite their close evolutionary relationship. Repeat motifs are easy to generate during cell division and DNA replication, thus can accrue fairly rapidly. Rather than being ‘junk’, the utility of DNA sequences that – at first glance – seem to nonsensically inflate genome size may have crucial utility after all.
Repeat DNA sequence motifs can also occur within chromosomes and play a role in the regulation of gene function (transcription (into RNA) and translation of a gene sequence into part of the working molecular machinery of the living cell (proteins or edited RNAs)). Genomes are no longer thought of as static ‘blueprints’ that the cellular machinery reads to build a body, rigidly following each step until the final project (a living organism) is complete. No, genomes are more like ‘choose-your-own-adventure’ books, whereby the cell can make different choices depending upon the environment that the cell, tissue or whole organism is facing at the time. That is why identical twins are never actually quite identical (although they might look it) and develop individual quirks and personalities. In genetics parlance, genes can be turned off or on, moved from place to place within the chromosome or between chromosomes, and individual genes can be invoked to make more than one product when it is read in a particular way or orientation (transcribed) or edited after being read (editing and or splicing of the gene transcripts). The science of transcriptomics (analysing the transcriptional products of the genome and patterns of gene activity) provides us with a looking glass into how genomes, and by extension organisms themselves, function. This science is of particular use in ecology when we want to look at the impacts of environmental influences on organismal biology and their responses to environmental variation, such as climate change. The tussock cottongrass ecotypes have been visited by functional ecologists to determine if their genomes are labile enough to show differences in gene expression between southerly and northerly populations, indicating that these plants can change plans with what genes to use, turn off or edit, depending upon their ecological circumstances.
In one example, whole genome transcriptomics have been used to determine the physiological responses of tussock cottongrass ecotypes to an extreme heat event (26.6 degrees centigrade, 2nd highest summer temperature then recorded). In this study employing whole genome transcriptomics (whereby every possible unique genetic transcript being expressed at the time of sampling is sequenced (“unigenes”) – here, total RNA is back converted to DNA and then sequenced using genomic sequencing technology), an astounding 423 thousand unique RNAs were identified, of which about 1500 (a mere 0.35 %) were being turned up, down or off. Some genes associated with the physiological response to heat were the ones that exhibited the greatest degree of change, including genes involved in response to dehydration and in cellular senescence. Different ecotypes responded significantly at variance to one another only when faced with extreme heat, and in one population in the extreme north, responded more adequately than southern ecotypes with more experience of such temperatures, suggesting that the capacity to respond to climate change may already have a foothold, at least in Alaska.
If the genome contains the answers to how well an organism might function in a dangerous future, the cottongrass genome is one of cautious optimism for its future survival.
© Arctic Institute of North America, University of Calgary
Site & Image Information

